










[儌僨儖憯椶僋儔儈僪儌僫僗]
[CO2擹弅婡峔偲廳扽巁僀僆儞桝憲懱偺敪尒]
[憯椶偺怴婏側帀幙拁愊惂屼場巕偺敪尒]
[嵟嬤偺廋巑榑暥偺僞僀僩儖]
儌僨儖椢憯僋儔儈僪儌僫僗
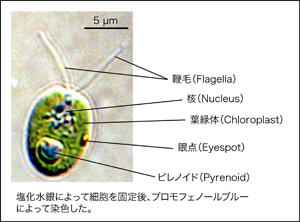
丂変乆偼偙傟傑偱偵CCM偺惂屼場巕乮Fukuzawa et al. PNAS 2001, Yoshioka et al. Plant Cell 2004乯丄柍婡扽慺偺桝憲偵娭傢傞僩儔儞僗億乕僞乕乮Yamano et al. PNAS 2015, Ohnishi et al. Plant Cell 2010乯丄塰梴寚朢忦審壓偱TAG偺拁愊傪惓偵惂屼偟丄拏慺寚朢壓偺岝崌惉妶惈偺梷惂偵娭梌偡傞怴婯側挷愡場巕乮Kajikawa et al. Plant Phys. 2015乯側偳傪摨掕偟偰偒傑偟偨丅傑偨丄cDNA忣曬傪棙梡偟偨僩儔儞僗僋儕僾僩乕儉夝愅偺庤朄傪妋棫偟乮Miura et al. Plant Phys. 2004乯丄僋儔儈僪儌僫僗偺慡僎僲儉夝撉傪曬崘偟傑偟偨乮Merchant et al. Science 2007乯丅
丂偝傜偵丄僋儔儈僪儌僫僗偺僎僲儉儔僀僽儔儕乕傗師悽戙僔乕働儞僒乕傪梡偄偨戝婯柾敪尰僾儘僼傽僀儖忣曬傪惍旛偟偰偍傝丄偙傟傜偺堚揱巕儕僜乕僗傪梡偄偰柧傜偐偵側偭偨懡偔偺壢妛揑抦尒偑榑暥偲偟偰敪昞偝傟偰偄傑偡丅僋儔儈僪儌僫僗偵偮偄偰偝傜偵徻嵶側忣曬傪抦傝偨偄曽偼丄僋儔儈僪儌僫僗偺僎僲儉夝撉偺榑暥傗丄壓婰偺擔杮岅偺憤愢婰帠傪嶲徠偟偰偔偩偝偄丅
-
擔杮岅憤愢乮僋儕僢僋偡傞偲PDF僼傽僀儖偑奐偒傑偡乯
- 暉郪廏嵠丄嶳栰棽巙丄妬愳徆岶
椢憯僋儔儈僪儌僫僗偵偍偗傞柍婡扽慺擹弅婡峔偲帀幙戙幱丂岝崌惉尋媶 22(3)乮2012) - 暉郪廏嵠丄媣曐梇徍丄嶳栰棽巙
椢憯僋儔儈僪儌僫僗偺僎僲儉偐傜怉暔偲摦暔偺婡擻傪扵傞 抈敀幙妀巁峺慺 53:1133-1143 (2008) - 暉郪廏嵠丄嶳栰棽巙
擇巁壔扽慺偵傛傞揮幨挷愡婡峔亅椢憯僋儔儈僪儌僫僗偺CO2擹弅婡峔 抈敀幙妀巁峺慺50:958-965 (2005)
CO2擹弅婡峔偲廳扽巁僀僆儞桝憲懱偺敪尒
丂悈拞偱偼CO2偼悈偲斀墳偟偰廳扽巁僀僆儞乮HCO3-乯偺宍偱懡偔懚嵼偡傞偙偲偑抦傜傟偰偄傑偡丅CO2偲堎側傝丄
揹壸傪懷傃偨廳扽巁僀僆儞偼嵶朎傗梩椢懱傪暍偆惗懱枌傪捠夁偡傞偙偲偑偱偒傑偣傫丅廬偭偰丄悈拞偺憯椶偼CO2傪嵶朎撪偵廫暘偵庢傝崬傓偙偲偑擄偟偔丄
岝崌惉偵晄棙側CO2寚朢娐嫬偵偝傜偝傟傑偡丅偙偺傛偆側娐嫬偵偍偄偰傕岝崌惉傪堐帩偟惗懚偡傞偨傔偵丄憯椶偼嵶朎枌偲梩椢懱曪枌偲偄偆2偮偺忈暻傪忔傝墇偊偰愊嬌揑偵廳扽巁僀僆儞傪庢傝崬傒丄
嵶朎撪偵擹弅偡傞偙偲偱岝崌惉傪峴偆偲峫偊傜傟偰偄傑偡丅偙傟偼僔傾僲僶僋僥儕傾傗憯椶偑丄岝僄僱儖僊乕傪梡偄偰嵶朎奜偐傜擻摦揑偵柍婡扽慺傪嵶朎撪偵桝憲偟丄
Rubisco廃曈偵CO2傪擹弅偡傞僔僗僥儉偵傛傞偙偲偑抦傜傟偰偍傝丄CO2擹弅婡峔乮CO2-concentrating mechanism: CCM乯偲柤晅偗傜傟偰偄傑偡丅
C4岝崌惉偺傛偆側堐娗懇忊嵶朎偲梩擏嵶朎偺暋嶨側憡屳嶌梡偵傛傞桳婡巁偺扙扽巁斀墳傪夘偡傞擹弅宱楬偲偼堎側傝丄CCM偼嵶朎枌傗梩椢懱枌偵嬊嵼偡傞桝憲懱傪夘偟偨柍婡扽慺偺捈愙揑側桝憲偵傛傞偲峫偊傜傟偰偄傑偡丅
憯椶偑偙偺傛偆側CO2擹弅宯(廳扽巁僀僆儞桝憲宯)傪帩偮偙偲偼1980擭偵惗棟妛揑偵敪尒偝傟傑偟偨偑
乮Badger et al. Plant Phys. 1980乯丄偦偺桝憲傪扴偭偰偄傞暘巕偺徻嵶偼挿傜偔晄柧偱偁傝丄
憯椶偺岝崌惉偺巇慻傒傪夝柧偡傞偆偊偱旕忢偵廳梫側栤戣偲偟偰巆偝傟偰偄傑偟偨丅
![]()
丂抧媴偺戝婥偵懡検偵娷傑傟偰偄傞巁慺偼丄岝崌惉偵傛傝悈偑暘夝偝傟偰惗偠傑偟偨偑丄媡偵CO2偼岝崌惉偵傛偭偰屌掕偝傟懕偗偰偒傑偟偨丅 岝崌惉偵傛偭偰挿偄擭寧拁偊傜傟偰偒偨愇扽愇桘傪擱傗偡偙偲偱丄戝婥拞偺CO2擹搙偼400 ppm偵憹壛偟丄尰嵼偺抧媴娐嫬偼壏抔壔丒怘椘晄懌丒僄僱儖僊乕屚妷側偳偺條乆側栤戣傪書偊偰偄傑偡丅 偙傟偵懳偟偰丄憯椶偑帩偮堚揱巕傪棙梡丒夵曄偟丄岝崌惉偺擻椡傪嬌尷傑偱崅傔偨僗乕僷乕怉暔傪憂弌偡傞偙偲偱夝寛偟傛偆偲偡傞怴偨側帋傒偑悽奅揑側嫞憟偺側偐恑傔傜傟偰偄傑偡丅 杮尋媶偱柧傜偐偵側偭偨憯椶偺廳扽巁僀僆儞桝憲僔僗僥儉傪僀僱傗僐儉僊側偳偺庡梫嶌暔偵摫擖偡傞偙偲偱CO2偺媧廂検偲惗嶻検傪崅傔丄忋婰偺栤戣傪夝寛偡傞偨傔偺僽儗僀僋僗儖乕偵偮側偑傞偙偲偑婜懸偝傟傑偡丅
-
娭楢偡傞榑暥偲憤愢
- Toyokawa C, Yamano T, Fukuzawa H
Pyrenoid Starch Sheath Is Required for LCIB Localization and the CO2-Concentrating Mechanism in Green Algae. Plant Physiology 182(4):1883-1893 (2020)


- Wang L, Yamano T, Takane T, Niikawa Y, Toyokawa C, Ozawa S, Tokutsu R, Takahashi Y, Minagawa J, Kanesaki Y, Yoshikawa H, Fukuzawa H
Chloroplast-mediated regulation of CO2-concentrating mechanism by Ca2+-binding protein CAS in the green alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 113(44):12586-12591 (2016)

- Yamano T, Sato E, Iguchi H, Fukuda Y, Fukuzawa H
Characterization of cooperative bicarbonate uptake into chloroplast stroma in the green alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 112(23):7315-7320 (2015)
- 嶳栰 棽巙丄暉郪 廏嵠
乽椢憯僋儔儈僪儌僫僗偵偍偗傞岝崌惉僞乕儃僄儞僕儞偺嬱摦偲惂屼乿
岝崌惉尋媶 29(1):14-28 (2019)
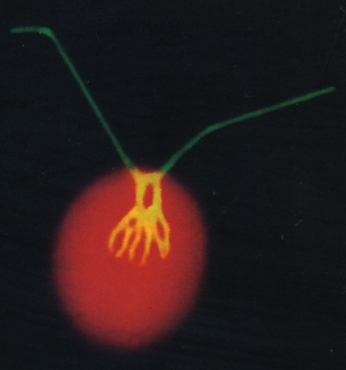
憯椶偺怴婏側帀幙拁愊惂屼場巕偺敪尒
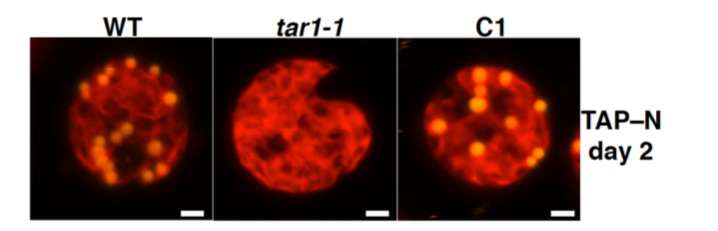
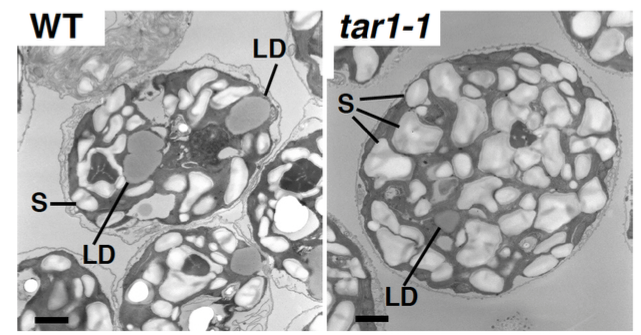
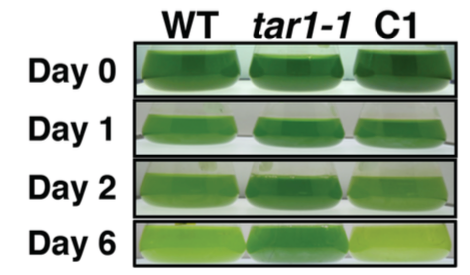
丂偙傟傑偱偵丄帀幙偺愼怓寀岝嫮搙傪巜昗偲偟偰丄僙儖僜乕僞乕暘庢僔僗僥儉傪梡偄偨曄堎懱偺僗僋儕乕僯儞僌傪峴偄丄拏慺寚朢忬懺偱僩儕傾僔儖僌儕僙儘乕儖乮TAG乯偺拁愊偑堎忢偵側傞曄堎懱 tag accumulation regulator1 (tar1)傪庢摼偟傑偟偨(Kajikawa et al. Plant Phys. 2015)丅曄堎懱tar1 偼丄恷巁攟抧傪梡偄偨岝廬懏塰梴忦審偱嵶朎傪拏慺婹夓忬懺偵偡傞偲丄栰惗宆偵斾傋偰(1) 僨儞僾儞偼拁愊偡傞偑 TAG 拁愊儗儀儖偑10 暘偺1偵掅壓偟丄(2) 嵶朎暘楐偑掆巭偟嵶朎僒僀僘偑戝偒偔側傝丄(3) 僋儘儘僼傿儖偺暘夝偑抶傟偰岝崌惉偺掆巭偑抶傟傞丄偲偄偆昞尰宆傪帵偟傑偟偨丅偙傟傜偺寢壥偐傜丄TAR1偼拏慺婹夓偵墳摎偟偨嵶朎暘楐丒岝崌惉丒帀幙拁愊偺惂屼偵學傢傞帠偑帵嵈偝傟傑偟偨丅TAR1堚揱巕偑僐乕僪偡傞僞儞僷僋幙偼丄峺曣偺Yak1 偲憡摨惈傪傕偮僞儞僷僋幙儕儞巁壔峺慺Dual-Specificity Tyrosine Phosphorylation-Regulated Kinase (DYRK)偺僒僽僼傽儈儕乕偺1偮偱偟偨丅偝傜偵丄嵶朎傪岝撈棫塰梴忦審偱拏慺寚朢偵偝傜偡偲丄僋儘儘僼傿儖傪堐帩偟偨傑傑丄嵶朎偁偨傝偺僨儞僾儞偵壛偊偰TAG偺拁愊検偑栰惗宆傛傝憹壛偡傞偲偄偆旕忢偵嫽枴怺偄昞尰宆傕娤嶡偝傟偰偄傑偡(榑暥弨旛拞)丅傑偨丄岝撈棫塰梴偐偮拏慺寚朢忦審偱栰惗姅偲tar1 曄堎姅偵偍偗傞堚揱巕敪尰傪RNA-seq偱栐梾揑偵夝愅偡傞偙偲偱丄tar1 曄堎姅偲栰惗姅偲偺娫偱敪尰儗儀儖偺嵎偑擣傔傜傟傞堚揱巕傪慖傃弌偟丄僞儞僷僋幙儕儞巁壔僾儘僥僆乕儉夝愅偺寢壥偲徠傜偟崌傢偣傞偙偲偱丄TAR1偑夘嵼偡傞TAG傗僨儞僾儞偺戙幱惂屼宯傪棟夝偡傞帠傪栚巜偟偰偄傑偡丅
-
娭楢偡傞榑暥摍
- Kajikawa M, Sawaragi Y, Shinkawa H, Yamano T, Ando A, Kato M, Hirono M, Sato N, Fukuzawa H
Algal dual-specificity tyrosine phosphorylation-regulated kinase, triacylglycerol accumulation regulator1, regulates accumulation of triacylglycerol in nitrogen or sulfur deficiency. Plant Physiol. 168(2):752-64 (2015)
嵟嬤偺廋巑丒攷巑榑暥偺僞僀僩儖
2020擭搙
[廋巑壽掱]丒椢憯偺C/N僗僩儗僗墳摎偵偍偗傞俀偮偺DYRK宆僉僫乕僛偺娭楢惈偲怴婏惂屼場巕
丒椢憯偵偍偗傞廳扽巁僀僆儞僠儍僱儖LCIA偺敪尰挷愡曄堎姅偺夝愅
丒Regulation of LCIA in Chlamydomonas reinhardtii and low sodium-dependency of photosynthesis in the diatom Chaetoceros gracilis
乮尋媶寢壥偺堦晹偑Marine Biotechnology帍乮2021乯偵宖嵹偝傟傑偟偨丅乯
2019擭搙
[攷巑壽掱]丒椢憯偺CO2擹弅婡峔偵偍偗傞僺儗僲僀僪僨儞僾儞忊偺栶妱
乮尋媶惉壥偑昅摢挊幰偲偟偰Plant Physiology帍乮2020乯偵丄傑偨嫟挊幰偲偟偰Plant Physiology帍乮2021乯偵宖嵹偝傟傑偟偨丅乯
丒扽慺偲拏慺偺塰梴僶儔儞僗墳摎偵偍偗傞椢憯偺僞儞僷僋幙儕儞巁壔峺慺TAR1偺婡擻
乮尋媶惉壥偑昅摢挊幰偲偟偰Plant&Cell Physiology帍乮2019乯偵宖嵹偝傟傑偟偨丅乯
[廋巑壽掱]
丒椢憯偺僺儗僲僀僪宍惉偵娭傢傞SAGA1僞儞僷僋幙偺CO2擹弅婡峔偵偍偗傞栶妱
丒C/N僗僩儗僗忦審偱僩儕傾僔儖僌儕僙儘乕儖傪崅拁愊偡傞椢憯偺CCDC124曄堎姅偺夝愅
2018擭搙
[廋巑壽掱]丒椢憯偵偍偗傞柍婡扽慺擹弅婡峔昁恵場巕LCIB偺嬊嵼惂屼
乮尋媶惉壥偺堦晹偑Plant Physiology帍乮2021乯偵宖嵹偝傟傑偟偨丅乯
丒椢憯偺C/N僗僩儗僗墳摎堎忢曄堎姅pp2c3偺夝愅
丒宂憯偵偍偗傞柍婡扽慺擹弅婡峔娭楢場巕偺扵嶕偲廳扽巁僀僆儞桝憲懱SLC4-1偺夝愅乮廋巑壽掱乯
乮尋媶惉壥偺堦晹偑Marine Biotechnology帍乮2021乯偵宖嵹偝傟傑偟偨丅乯
丒宂憯偺帀幙戙幱夵曄偵傛傞儕僔僲乕儖巁惗嶻擻偺夵椙
2016擭搙
[攷巑壽掱]丒A calcium-binding protein CAS regulates the CO2-concentrating mechanism in the green alga Chlamydomonas reinhardtii
乮尋媶惉壥偑昅摢挊幰偲偟偰PNAS帍乮2016乯偵宖嵹偝傟傑偟偨丅乯
[廋巑壽掱]
丒宂憯Chaetoceros gracilis偵偍偗傞儕僔僲乕儖巁偺惗嶻
乮尋媶惉壥偺堦晹偑Scientific Reports帍乮2016乯偵宖嵹偝傟傑偟偨丅乯
丒CO2擹弅婡峔傪惂屼偡傞CCM1暋崌懱偍傛傃CAS偺夝愅
乮尋媶惉壥偺堦晹偑PNAS帍乮2016乯偵宖嵹偝傟傑偟偨丅乯
丒椢憯僋儔儈僪儌僫僗偲宂憯僣僲働僀僜僂偵偍偗傞廳扽巁僀僆儞桝憲懱偺夝愅
乮尋媶惉壥偺堦晹偑PNAS帍乮2015乯偵宖嵹偝傟傑偟偨丅乯
丒僋儔儈僪儌僫僗拏慺寚朢墳摎曄堎姅偍傛傃僔儘僀僰僫僘僫AtYak1曄堎姅偺夝愅
2015擭搙
[廋巑壽掱]丒僩儕傾僔儖僌儕僙儘乕儖拁愊堎忢姅tar1偺扨棧偲夝愅
乮尋媶惉壥偺堦晹偑Plant Physiology帍乮2015乯偵宖嵹偝傟傑偟偨丅乯
丒柍婡扽慺擹弅擻傪惂屼偡傞怴婯場巕CrCAS偺婡擻夝愅
乮尋媶惉壥偺堦晹偑PNAS帍乮2016乯偵宖嵹偝傟傑偟偨丅乯
2014擭搙
[廋巑壽掱]丒椢憯僋儔儈僪儌僫僗偵偍偗傞僗僋傾儗儞戙幱偺惂屼婡峔偺夝愅
乮尋媶惉壥偺堦晹偑PLOS ONE帍乮2015乯偵宖嵹偝傟傑偟偨丅乯
丒椢憯僋儔儈僪儌僫僗偵偍偗傞掅CO2桿摫惈枌僞儞僷僋幙LCIA偲HLA3偺婡擻夝愅
乮尋媶惉壥偺堦晹偑PNAS帍乮2015乯偵宖嵹偝傟傑偟偨丅乯
丒椢憯僋儔儈僪儌僫僗栰惗姅C9偺SNPs夝愅偍傛傃宂憯僉乕僩僙儘僗偺CO2寚朢忦審壓偵偍偗傞岝崌惉摿惈
丒椢憯僋儔儈僪儌僫僗偵偍偗傞CO2墳摎偵堎忢傪帩偮曄堎姅偺扨棧偍傛傃僆乕僩僼傽僕乕娭楢堚揱巕偺婡擻夝愅
2013擭搙
[廋巑壽掱]丒岝偲CO2偵埶懚偟偰嬊嵼曄壔偡傞梩椢懱僞儞僷僋幙LCIB偺婡擻夝愅
乮尋媶惉壥偺堦晹偑Photosynthesis Research帍乮2014乯偵宖嵹偝傟傑偟偨丅乯
丒儐僯乕僋側摐寢崌儌僕儏乕儖傪娷傓價僼傿僘僗嬠偺兛-galactosidase偼B宆寣塼宆峈尨偵摿堎揑偵嶌梡偡傞
乮尋媶惉壥偑昅摢挊幰偲偟偰Glycobiology帍偵宖嵹偝傟傑偟偨丅乯
丒Bifidobacterium longum subsp.longum JCM1217桼棃GH43僌儕僐僔僟乕僛偺夝愅
丒慄拵Caenorhabditis elegans傊偺僌儖僐僒儈儞搳梌偵傛傞庻柦傊偺塭嬁偺夝愅
丒價僼傿僘僗嬠Bifidobacterium longum subsp. longum JCM 1217桼棃GH31峺慺偺夝愅
丒椢憯僋儔儈僪儌僫僗偵偍偗傞僆儗僀儞巁戙幱偺夵曄側傜傃偵僆乕僩僼傽僕乕偺婡擻夝愅